
|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| Thread ▼ Details |
|
Thread Info
|
|
|
| Author | Topic: Rebuttal To Creationists - "Since We Can't Directly Observe Evolution..." | |||||||||||||||||||||||||||||||||||||||
|
Taq Member  Posts: 10085 Joined: Member Rating: 5.6
|
Kleinman writes: You are so stupid. The Kishony experiment works quickly because his lineages can accumulate their 1/(mutation rate) replications without having to deal with biological competition. You didn't address what I said. Here it is again. The Lenski experiment had a constant environment for 10's of thousands of generations. Are you saying that a constant environment does not disqualify an experiment? As to the Kishony experiment, what would the dynamics of adaption to chloramphenicol look like if the E. coli in the experiment were undergoing sexual recombination? How would it compare to a population of asexual E. coli adapting to the same environment? Imagine if scientists published a paper on that very experiment? Well we don't have to imagine.
quote: Look at the title of that paper. Wouldn't you know it, sexual recombination speeds up adaptation, and it does so by alleviating clonal inteference. More from the paper:
quote: CM tolerance is chloramphenicol resistance. Sexual recombination combines different beneficial mutations in different genes into the same genetic background, thereby speeding up adaptation to chloramphenicol.
The only reason that Desai's population doesn't see the effect of clonal interference is that he lets his population increase the frequencies of a few adaptive alleles and then induces sexual reproduction after 90 generations of constant selection. Yes, it is sexual recombination that alleviates clonal interference.
If Desai allowed his environment to change, who knows if any alleles would have increased in frequencies and who knows what kind of recombination events would have happened. Why wouldn't those same beneficial mutations increase in frequency if there was sexual recombination in every generation?
Why don't you do the math with a varying environment and see how much recombination helps? The experiment was already run. It helps.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
The Peabody et. al. (2017) paper mentioned earlier has a great illustration of how adaptation differs in asexual and sexual populations:
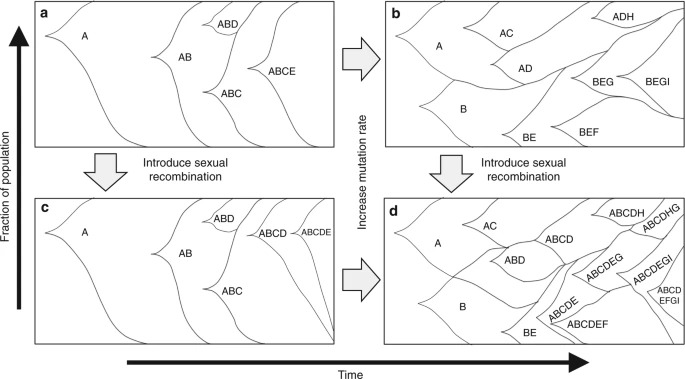 from the paper: Theoretical impacts of mutation rate and sexual recombination on population structures. a Mutationally limited fitness landscape, where most mutations fix in a single sweep with very little inter-clonal competition. b Increased mutation rate over (a), more competition and faster evolution observed, but some mutations are lost due to inter-clonal competition. c Adding sexual recombination to (a), due to few available beneficial mutations, no strong influence on evolution is observed. d Both sexual recombination and increase in mutation rate from (a), more rapid evolution is observed and fewer beneficial mutations are lost due to clonal interference The two variables here are mutation rate and recombination. The two plots on the left are with the lower mutation rate. The two plots at the bottom are with sexual recombination. a. low mutation, asexualb. low mutation, sexual c. high mutation, asexual d. high mutation, sexual Even at low mutation rates, you can see that the sexual population gets all 5 beneficial mutations which the asexual population is limited to 4 because clonal interference drove the ABD lineage to extinction. In the sexual population, recombination between the ABC and ABD lineages put all 5 beneficial mutations into the same background. When mutation rates are even higher, the differences can be even more stark.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6
|
Kleinman writes: If you understood the Kishony or Lenski experiments, you would have done the mathematics long ago, but you don't. You still haven't addressed what I said. The Lenski experiment had a constant environment for 10's of thousands of generations. Are you saying that a constant environment does not disqualify an experiment? The only way that Desai could get his experiment to work was to use a constant environment, let adaptive alleles increase in frequencies for 90 generations in a constant environment, and then induce sexual reproduction. The adaptive alleles would increase in frequency if there was sexual reproduction in every generation.
You are so stupid, you think that recombination works the same way in a varying environments. How does recombination work differently in varying environments?
You stupidly extrapolate a constant environment to all environments. The Lenski experiment used a constant environment of 10's of thousands of generations. "The Kishony and Lenski experiments demonstrate exactly how descent with modification operates."--Kleinman
quote: Yes, it does.
quote: And you are so stupid, you extrapolate this result to all evolution. So says the person who extrapolates the Kishony/Lenski experiment to all of evolution.
Because they aren't increasing in frequency. The adaptive alleles change as the environment changes, moron. You are really mixed up now. You were saying that Desai had to asexual reproduction in a constant environment in order to increase the frequency of beneficial alleles. Why wouldn't those same alleles increase in fitness with sexual reproduction in every generation and a constant environment?
Why did Desai use 90 generations of constant selection? Why did Lenski use 10's of thousands of generations of constant selection?
Yeah, you simpleton, in a constant environment you get a slight increase in relative fitness. Now, learn that this principle doesn't apply when the environment and selection conditions are changing. Why don't you show us the principle. Show us a real world example that compares asexual and sexual reproduction in a changing environment. Show us the math. Put up or shut up.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
Kleinman writes: The problem is their theory doesn't match up with reality, Except that it does. Peabody et al. observed those same effects in their experiment.
Sexual recombination and increased mutation rate expedite evolution of Escherichia coli in varied fitness landscapes Beneficial mutations for CM resistance can occur in different genes which makes it a candidate for studies on sexual recombination. They found that the combination of a faster mutation rate and sexual recombination increased the speed of adaptation to CM resistance.  The genderless are the sexual reproducers, and higher mutation rates were induced with arabinose (ara). Therefore, the sexual reproducers with high mutation rates are the genderless ara+, and they adapted to CM the fastest.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
The more you look around the more examples you can find of sexual recombination increasing the efficiency of adaptation.
quote: The rec+ populations were recombination positive. Wouldn't you know it, recombination alleviated clonal interference and sped up adaptation by putting beneficial mutations from different lineages into the same genetic background. Seems to be a repeating theme.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
Kleinman writes: Is that all you can find is constant environment experiments with a single selection pressure. The Lenski experiment used a constant environment and a single selection pressure for 50,000 generations. "The Kishony and Lenski experiments demonstrate exactly how descent with modification operates."--Kleinman
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6
|
Kleinman writes: I've said it many times, you just don't get it. It takes 1/(mutation rate) replications in a single selection pressure environment to get an adaptive mutation. Joshua and Esther Lederberg published a hallmark paper in 1951 titled, "Replica Plating and Indirect Selection of Bacterial Mutants", which can be found here: REPLICA PLATING AND INDIRECT SELECTION OF BACTERIAL MUTANTS - PMC Luria and Delbruck went on to explain the processes that undergirded the Lederberg's results which later won Luria and Delbruck a Nobel Prize. There is one interesting observation in the Lederberg paper: "The culutre is fully sensitive to the phage T-1, as well as to streptomycin, and like most E. coli strains gives rise to resistant mutants at rates of approximately 10E-7 and 10E-10 per division, respectively." In this example we saw a beneficial mutation rate of 1 in 10 million and 1 in 10 billion to two different selection pressures using the same strain of E. coli. Kleinman is telling us that one beneficial adaptation every billion divisions is some universal constant, or something of the like. It is so universal that it can even be applied to human evolution. However, in another experiment using E. coli we see beneficial mutation rates that are quite different than what Kleinman claims. If Kleinman's math can't even apply universally to evolution in E. coli, what hope does it have of applying to any other species?
When recombination is a possibility, a changing environment reduces the probability because the selection condition is changing therefore, the adaptive alleles are changing. Reduces the probability of what, and compared to what? Would a changing environment cause clonal interference in a sexually reproducing population? NO. Would beneficial alleles in the new environment increase in frequency in the new environment in sexually reproducing populations? YES Would you get a combination of beneficial alleles faster in a changing environment with a sexually reproducing population compared to an asexual population? YES
So you dream. None of the adaptive alleles would have had a chance to increase in frequency. In a constant environment, why wouldn't beneficial alleles increase in frequency in a sexually reproducing population?
You should know the answer to this one dummy, even Tany blundered into this one. I do know the answer. Sexual recombination doesn't change in a changing environment. It works the same way. When humans move to a different environment their gametes still work in the very same way.
The Lenski experiment would operate more slowly in a changing environment. You said the Lenski experiment demonstrates exactly how descent with modification works, and it had 50,000 generations in the same exact environment. You reject the Desai experiment because it had a constant environment for 90 generations. Hypocrisy much?
The genius virologist has just proven that 3 drug combination therapy does not work for the treatment of HIV. I'm not the one rejecting published science. You are.
Biological evolution only gets slower with changing environments and multiple selection pressures. You really don't know anything about biological evolution. Then show it in a real population. Show how an asexual and sexual population evolve in changing environments with multiple selection pressures.
I've given you many empirical examples, HIV, insects, and weeds. All of which increase their rate of adaptation through sexual recombination.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
quote: Wouldn't you know it, recombination can speed up the acquisition of multi-herbicide resistance.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
quote: Recombination is an important factor in evolving multidrug resistance in HIV.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
Kleinman writes: So you think that non-target site resistance is your answer? Do you even know what non-target site resistance is? That exists with bacteria as well. Why don't you tell us what non-target site resistance is? So your only response is to quiz people on terminology? That's it?
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
Kleinman writes: That's right, in a constant environment, single selection pressure environment it takes a billion replications for each adaptive mutation. Joshua and Esther Lederberg published a hallmark paper in 1951 titled, "Replica Plating and Indirect Selection of Bacterial Mutants", which can be found here: REPLICA PLATING AND INDIRECT SELECTION OF BACTERIAL MUTANTS - PMC Luria and Delbruck went on to explain the processes that undergirded the Lederberg's results which later won Luria and Delbruck a Nobel Prize. There is one interesting observation in the Lederberg paper: "The culutre is fully sensitive to the phage T-1, as well as to streptomycin, and like most E. coli strains gives rise to resistant mutants at rates of approximately 10E-7 and 10E-10 per division, respectively." In this example we saw a beneficial mutation rate of 1 in 10 million and 1 in 10 billion to two different selection pressures using the same strain of E. coli. Kleinman is telling us that one beneficial adaptation every billion divisions is some universal constant, or something of the like. It is so universal that it can even be applied to human evolution. However, in another experiment using E. coli we see beneficial mutation rates that are quite different than what Kleinman claims. If Kleinman's math can't even apply universally to evolution in E. coli, what hope does it have of applying to any other species?
Kleinman writes: When there are multiple selection pressures and the environment is changing, it takes exponentially more replications to adapt. It isn't exponential in sexually reproducing populations. Haven't you been paying attention? Beneficial alleles for each of those challenges can be combined into a single genetic background through recombination. You still don't understand how sexual reproduction works.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
Kleinman writes: They might as well forget combination therapy for treating HIV, Taq says it will fail. You are a brilliant virologist. He is also an expert in non-target site resistance even though he doesn't know what it is. I am citing published science which demonstrates recombination is an important factor in evolving multidrug resistance.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
Kleinman writes: Ah, so it was a phage that evolved a human, that's brilliant. Tell us all about it. Still no response to the actual material???? Joshua and Esther Lederberg published a hallmark paper in 1951 titled, "Replica Plating and Indirect Selection of Bacterial Mutants", which can be found here: REPLICA PLATING AND INDIRECT SELECTION OF BACTERIAL MUTANTS - PMC Luria and Delbruck went on to explain the processes that undergirded the Lederberg's results which later won Luria and Delbruck a Nobel Prize. There is one interesting observation in the Lederberg paper: "The culutre is fully sensitive to the phage T-1, as well as to streptomycin, and like most E. coli strains gives rise to resistant mutants at rates of approximately 10E-7 and 10E-10 per division, respectively." In this example we saw a beneficial mutation rate of 1 in 10 million and 1 in 10 billion to two different selection pressures using the same strain of E. coli. Kleinman is telling us that one beneficial adaptation every billion divisions is some universal constant, or something of the like. It is so universal that it can even be applied to human evolution. However, in another experiment using E. coli we see beneficial mutation rates that are quite different than what Kleinman claims. If Kleinman's math can't even apply universally to evolution in E. coli, what hope does it have of applying to any other species?
The probability of an adaptive recombination event occurring, slow poke. You can speculate otherwise but combination therapy for HIV, weeds, and insects says you are wrong, they work. I'm not speculating. The scientific literature demonstrates that recombination is an important factor in multidrug resistance in HIV and multi-herbicide resistance in weeds.
Even Tany knows that the most fit variant changes as the environment changes. But now I understand why you have this preoccupation with a constant environment, single selection pressure systems, it is the limit of your understanding. The real world isn't made of constant environments with a single selection pressure. You have very limited education. Apparently, you haven't been reading the papers I have cited for you.
quote: Sexual recombination allows multiple beneficial alleles to move towards fixation simultaneously. You keep forgetting that.
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6
|
Kleinman writes: This is your link. If you think it proves UCD, go for it. I think it proves your claim about 1 adaption per 1 billion replications wrong. Joshua and Esther Lederberg published a hallmark paper in 1951 titled, "Replica Plating and Indirect Selection of Bacterial Mutants", which can be found here: REPLICA PLATING AND INDIRECT SELECTION OF BACTERIAL MUTANTS - PMC Luria and Delbruck went on to explain the processes that undergirded the Lederberg's results which later won Luria and Delbruck a Nobel Prize. There is one interesting observation in the Lederberg paper: "The culutre is fully sensitive to the phage T-1, as well as to streptomycin, and like most E. coli strains gives rise to resistant mutants at rates of approximately 10E-7 and 10E-10 per division, respectively." In this example we saw a beneficial mutation rate of 1 in 10 million and 1 in 10 billion to two different selection pressures using the same strain of E. coli. Kleinman is telling us that one beneficial adaptation every billion divisions is some universal constant, or something of the like. It is so universal that it can even be applied to human evolution. However, in another experiment using E. coli we see beneficial mutation rates that are quite different than what Kleinman claims. If Kleinman's math can't even apply universally to evolution in E. coli, what hope does it have of applying to any other species?
You should publish your finding that combination treatment doesn't work for HIV, and combination therapy doesn't work for weeds and insects because you are so smart. IT'S ALREADY PUBLISHED!!!
|
|||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.6 |
Kleinman writes: You think this causes UCD, so go for it, explain how a phage makes this possible. I think it proves your claim about 1 adaption per 1 billion replications wrong. You still can't respond to this???? Joshua and Esther Lederberg published a hallmark paper in 1951 titled, "Replica Plating and Indirect Selection of Bacterial Mutants", which can be found here: REPLICA PLATING AND INDIRECT SELECTION OF BACTERIAL MUTANTS - PMC Luria and Delbruck went on to explain the processes that undergirded the Lederberg's results which later won Luria and Delbruck a Nobel Prize. There is one interesting observation in the Lederberg paper: "The culutre is fully sensitive to the phage T-1, as well as to streptomycin, and like most E. coli strains gives rise to resistant mutants at rates of approximately 10E-7 and 10E-10 per division, respectively." In this example we saw a beneficial mutation rate of 1 in 10 million and 1 in 10 billion to two different selection pressures using the same strain of E. coli. Kleinman is telling us that one beneficial adaptation every billion divisions is some universal constant, or something of the like. It is so universal that it can even be applied to human evolution. However, in another experiment using E. coli we see beneficial mutation rates that are quite different than what Kleinman claims. If Kleinman's math can't even apply universally to evolution in E. coli, what hope does it have of applying to any other species?
You think that combination therapy doesn't work because of recombination, there is far more literature that says it does work. It works for HIV, weeds, and insects. You are wrong. I cited the scientific literature which clearly stated that recombination is an important factor in multidrug resistance in HIV. Ignore it all you want, it's still there.
Show me the paper that explains how drug resistance evolves. I already did that. Sexual recombination and increased mutation rate expedite evolution of Escherichia coli in varied fitness landscapes | Nature Communications
Only in a single selection pressure constant environment and that only gives a slight improvement in fitness. In a multi-selection pressure environment with multiple selection pressures, it's a different game even if you think that evolution works the same in a constant single-pressure environment as in a varying multiselection pressure environment. quote:
|
|||||||||||||||||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024
 (1)
(1)
