|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| Thread ▼ Details |
|
Thread Info
|
|
|
| Author | Topic: Rebuttal To Creationists - "Since We Can't Directly Observe Evolution..." | ||||||||||||||||||||||||||||||||||||||||
|
Taq Member  Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes: And the selection condition doesn't change the math, it only changes the target gene(s) as demonstrated by the different selection conditions use in the Kishony and Lenski experiments. Sexual selection does change your math. All of your math is based on one beneficial mutation reaching fixation at a time. That's ridiculous. That is not how it works in sexually reproducing populations.
There are not many selection conditions on a savanna, but a few, are starvation, dehydration, thermal stress (both excessively high and excessively low temperatures), disease (bacterial, fungal, parasitic, viral), toxins, and predation, to list a few. It would be sad for some member of the population to get an adaptive mutation that would give a step toward standing upright and end up dying of tetanus or starvation or any of the myriad of other selection conditions that member would face. There wouldn't be much of an improvement in fitness from that first mutation. The earliest populations wouldn't have to be fully adapted to the open savanna. They could live at the edges of their arboreal range and only venture out a bit into the savanna. As they acquired more an more adaptations to the savanna they could move further out into the savanna which would remove them from competition with other arboreal apes. This is nothing like antibiotic selection.
If both parents are homozygous for the resistance allele, then you are doing 2 random trials for the next adaptive mutation in that replication.
Again, there is more than one gene in a genome. Parent One is homozygous for a beneficial mutation in gene A. Parent Two is homozygous for a beneficial mutation in gene B. All offspring will be heterozygous for both beneficial mutations. There's no need for another random trial to get the 2nd mutation in each parent.
Drug-resistant bacteria appear as a matter of course simply by neutral evolution. The same for phage resistance, as shown in both the Lederberg plate replica experiment and the Luria and Delbruck fluctuation assay. In those experiments, they start with a single bacterium and grow a population. They then expose that population to antibiotics or phage. They find resistant bacteria that already had the resistance mutation before being exposed to either challenge. However, as discussed in the Lederberg paper, antibiotic resistance appeared 1,000 times less often than phage resistance. How do you explain this difference in the appearance of resistance in both cases?
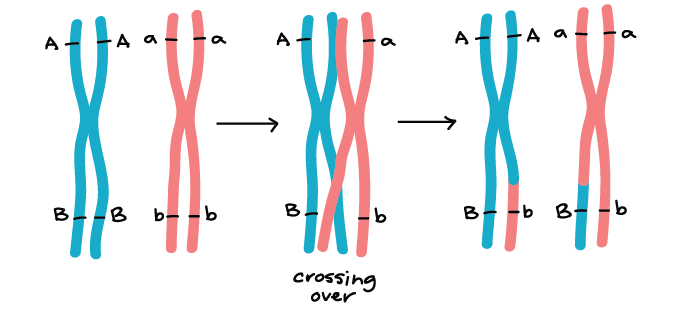
Doesn't relative fitness differences of different variants determine which variants increase in frequency and which decrease? That is only for alleles of the same gene. This is not true for different genes in a sexually reproducing population. Again, a genome has more than one gene. If there are two beneficial alleles for two different genes then they will both increase in frequency unless there is an interaction between the genes that lowers fitness. For example, the two mutations in different genes that I spoke about earlier, one for lighter skin and one for lactase persistence. They are both beneficial on their own, and they don't compete with one another because the mutations are on different genes. Eukaryotes have more than one chromosome. Genes on separate chromosomes are not linked in any way. They independently disperse through the population.Even then, meiosis will switch alleles across the paired chromosomes so you will have different linkages between alleles of different genes on the same chromosome through time. There is about 1 cross over event per chromosome per offspring, at least in humans. The only difference in the math is that for a clonal replicator, each genome replication is one adaptive allele replication. There is more than one gene in the genome.
This is the paper that explains how to compute the probability of adaptive mutations occurring at two or more genetic loci. Those are asexual populations under extremely stringent selection conditions, both of which are not true for human evolution.
And you still don't get it, competition and fixation slow adaptation. This calculation is all about determining the probability of getting an A1A2 variant as a function of the entire population size. YES, IN AN ASEXUAL POPULATION. What are you not getting here? Just to repeat . . . Parent One is homozygous for a beneficial mutation in gene A. Parent Two is homozygous for a beneficial mutation in gene B. All offspring will be heterozygous for both beneficial mutations. There's no need for another random trial to get the 2nd mutation in each parent.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1 |
Kleinman writes: All these alleles must first exist in the population for this process to work which means you need to start with billions of replications of the replicator (Finches in this case) before you even have a chance for this type of selection process to work. Then, when this process occurs, you reduce the diversity of the lineage that remains. Billions of replications would put beneficial mutations in many different genes. You keep asserting that those mutations on separate genes stay separate from one another. They don't.
You start with a diverse population of Finches with a diverse distribution of alleles that cause all different sizes and shapes of beaks. You put that population into an environment with a food source that requires a particular beak size and shape to feed, for example, insects in crevices. Those variants with wrong-shaped beaks die from starvation leaving a population with a high frequency of beaks that are long and narrow. Those remaining variants breed among themselves and any alleles that improve beak size for that food source have an increased probability of recombining into future offspring. That same process occurs for those variants with short, stout beaks that are suitable for eating nuts in some other environment. Ultimately, this process reduces the diversity of the population. Actually, hybridization was a big part of the process:
quote: This was an increase in genetic variation. It was the mixture of mutations from different lineages that resulted in adaptation, something you can't seem to get your head wrapped around.
You simply do not have sufficient population size to explain the evolution of humans and chimps from a common ancestor to what we see today using this selection process. The only reason you are saying this is because you only allow one beneficial mutation to move towards fixation at a time, which is ridiculous.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1 |
Kleinman writes: You still haven't learned that the competition and fixation process slows the DNA adaptive evolutionary process. And you haven't thought through what is required for recombination to operate. Parent One is homozygous for a beneficial mutation in gene A. Parent Two is homozygous for a beneficial mutation in gene B. All offspring will be heterozygous for both beneficial mutations. There's no need for another random trial to get the 2nd mutation in each parent. Please show me how this is wrong.
You are trying to simulate a savanna-like Kishony's experiment but you don't even realize it. Do you think that starvation, dehydration, thermal stress (both excessively high and excessively low temperatures), disease (bacterial, fungal, parasitic, viral), toxins, and predation don't exist on the edge of the savanna? Human ancestors weren't plucked from their arboreal environment and placed smack dab in the middle of the savanna. That's not what happened. That is what happens in the analogous Kishony experiment. The Kishony experiment requires adaptation in almost a single generation, something our human ancestors would not have had to do.
Are you ready to do the probability mathematics of random recombination? Here's another hint for you, it is a trinomial distribution. Parent One is homozygous for a beneficial mutation in gene A. Parent Two is homozygous for a beneficial mutation in gene B. All offspring will be heterozygous for both beneficial mutations. There's no need for another random trial to get the 2nd mutation in each parent. Please show me how this is wrong.
First, you explain how the phage got the resistance allele. The same way streptomycin gets antibiotic resistance. Do cancer drugs get drug resistance? It is the bacteria that are phage resistant. The phage aren't phage resistant. That makes no sense. The bacteria had already evolved phage resistance before they came into contact with phage, so it wasn't the phage that gave the bacteria a resistance allele. So I will ask again. As discussed in the Lederberg paper, antibiotic resistance appeared 1,000 times less often than phage resistance. How do you explain this difference in the appearance of resistance in both cases? Doesn't that same process occur with chimps as well? Why don't they have a population of over 7 billion today? Please address what I wrote: you: Doesn't relative fitness differences of different variants determine which variants increase in frequency and which decrease? me: That is only for alleles of the same gene. This is not true for different genes in a sexually reproducing population. Again, a genome has more than one gene. If there are two beneficial alleles for two different genes then they will both increase in frequency unless there is an interaction between the genes that lowers fitness. For example, the two mutations in different genes that I spoke about earlier, one for lighter skin and one for lactase persistence. They are both beneficial on their own, and they don't compete with one another because the mutations are on different genes. Eukaryotes have more than one chromosome. Genes on separate chromosomes are not linked in any way. They independently disperse through the population. Even then, meiosis will switch alleles across the paired chromosomes so you will have different linkages between alleles of different genes on the same chromosome through time. There is about 1 cross over event per chromosome per offspring, at least in humans. You still don't get or don't want to get that the only difference in the math between asexual replicators and sexually reproducing replicators is the former you can use genome replications while in the latter, use allele replications. There is more than one gene in the genome.
You are not taking into account the population sizes necessary and population recovery necessary for this kind of selection to occur more than once. You don't need population bottlenecks in order for beneficial mutations in different genes to all reach fixation in parallel.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes: Taq, you have mastered Mendelian Genetics. Now you need to master the mathematics of selection. What must happen to a population in the wild in order for your recombination example to have a reasonable probability of occurring? 1. Each of the two beneficial mutations must be under positive selection.2. Free interbreeding within the population. 3. Positive selection lasts long enough so that both beneficial mutations reach a small percentage of the population. If 5% of the population is heterozygous for one of the two beneficial mutations then 2.5% of births will have parents where each one has one of the two beneficial mutations. If they are heterozygous, 12.5% of their children will have both mutations. Dual carriers will only increase as each mutation increases in frequency until 100% of the population has both mutations if the benefice of each mutation is strong enough.
I'm not aware of any real environments devoid of selection pressures. I'm not aware of very many environments where only a few individuals survive out of billions in a single generation.
And the Kishony experiment requires about 30 generations of doubling for each adaptive mutational step. Again, they are asexual organisms. That's why there are steps. If they are reproducing sexually then multiple steps could occur in one set of 30 generations and be combined into a single genome.
You just don't want to accept the fact that this kind of selection bottlenecks a population. Bottlenecks do occur, they just aren't necessary.
Are you saying that phages are what give humans the reproductive advantage over chimps? I thought phages only infect bacteria. So I will ask again. As discussed in the Lederberg paper, antibiotic resistance appeared 1,000 times less often than phage resistance. How do you explain this difference in the appearance of resistance in both cases? Sure, there are thousands of coding genes and most of the genome controls the expression of the genes. Every gene and the rest of the genome are potential targets for mutations. What's the probability of more than one adaptive mutation occurring in a genome in a single replication In a single replication the odds are low. Across hundreds of millions of replications in humans, the odds are extremely high that there will be different beneficial mutations in a whole host of different genes. You want to make the claim that only the fittest mutation out of all them will move towards fixation at the demise of all those other mutations in other genes. THIS ISN'T TRUE. All of those mutations will have their frequencies changed by comparison to the other alleles for that gene, not the frequency of alleles in other genes.
That's an interesting claim. Since you are in speculation mode, give us an example of this in human evolution and explain why it didn't happen with chimps.
Speculation mode?????? Are you once again denying this scenario? Parent One is homozygous for a beneficial mutation in gene A. Parent Two is homozygous for a beneficial mutation in gene B. All offspring will be heterozygous for both beneficial mutations. There's no need for another random trial to get the 2nd mutation in each parent. Please show me how this is wrong. This would have happened in the chimp lineage as well. The difference is that the chimp lineage and the human lineage adapted to different environments, so they had different mutations reach fixation. It's one of the basic concepts in evolution.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes: Do you have any empirical examples that correlate with your calculation? It's simply math. If you agree that selection increases the frequency of beneficial alleles then the outcome I described is inevitable. And I already showed you the real world example:
quote: HIV doesn't do what you claim. HIV is diploid and does recombination. If you put human ancestors in an environment where only a few out of billions survived in each generation then they would go extinct. However, that is not what happened in the evolution of humans. It is the extremely harsh environment that makes your HIV model irrelevant to human evolution. I fully agree that if the founding population of the human lineage was put in a similar extreme environment that we would not be here today.
That happens all the time with antimicrobial therapy. But it didn't happen in human evolution.
The same kind of DNA adaptive evolutionary steps happens with sexually replicating organisms. The resistance alleles must form by a sequence of adaptive mutations accumulating on an allele. Parent One is homozygous for a beneficial mutation in gene A. Parent Two is homozygous for a beneficial mutation in gene B. All offspring will be heterozygous for both beneficial mutations. There's no need for another random trial to get the 2nd mutation in each parent. Please show me how this is wrong. I'm still trying to figure out how you can get two variants fixed in a population simultaneously. Are you remembering that they are on different genes? I'm guessing you forgot that part. Are you also forgetting about separate chromosomes and meiosis?
I've never studied the experiment, but if you think it explains how humans and chimpanzees evolved from a common ancestor, please explain. I am asking you to explain. So I will ask again. As discussed in the Lederberg paper, antibiotic resistance appeared 1,000 times less often than phage resistance. How do you explain this difference in the appearance of resistance in both cases? For a mutation rate of 1e-8, that probability is on the order of 1e-16. Where are the units on these rates?
I don't make any claims about the fixation of any beneficial alleles in humans. Bullshit. From message 246: "The math is way beyond you. 20 million beneficial mutations * 300 generations/fixation = 6 billion generations" You have one beneficial moving to fixation at a time, and no other beneficial mutations are even allowed to occur or move towards fixation while one beneficial mutation is moving towards fixation. That's ridiculous. This is ALL about how different mutations in different genes move towards fixation. All you can seem to do is see one gene and one beneficial mutation at a time. You simply can't wrap your head around the idea that there can be more than one possible beneficial mutation in more than one gene. You also can't seem to understand that these mutations are spread over many chromosomes.
How does a beneficial mutation occurring in an Alaskan Eskimo get recombined with a beneficial mutation from an Australian Aborigine? It may take a long time. The most recent history of humans has seen isolated subpopulations. This doesn't seem to be the case for most of our history where we were found in Africa.
With regards to your scenario, where's the beef? You pass your Mendelian Genetics exam. It demonstrates the very thing you claim can't happen, the combination of different beneficial mutations into a single genome.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes: Taq needs a little help with his math. A probability of 0.025 does not equal 25%, it equals 2.5%. And according to his calculation, that's when 5% of his population has the A allele and another 5% have the B allele. That is 1000 members each in his 20,000-member population. How many generations does it take for these beneficial alleles to "fix" and this recombination process to occur? I don't need any help. We have two genes with two alleles, Aa and Bb. Parent A is Aa bb, Parent B is aa Bb. Half of the children will be Aa and half will be Bb. The odds of being Aa and Bb are the product of those probabilities, 0.25 or 25%.
Define the following variables: n – is the total population size. nA – is the number of members in the population with beneficial allele A. nB – is the number of members in the population with beneficial allele B. nC – is the number of members in the population that have neither beneficial allele A nor beneficial allele B. In addition, we have the following condition: nA + nB + nC = n. And the frequency of each of the variants are: f_A = nA/n f_B = nB/n f_C = nC/n You are calculating 3 alleles at the same gene, not 3 mutations in 3 different genes. The frequency of each mutation can be 90%, and that obviously does not add up to the total population size. In other words, you are modeling AA, Aa, and aa. You are not modeling the combination of Aa and Bb.
Perhaps Taq wants to try the example of more than 2 adaptive alleles? Perhaps you should try modeling mutations in different genes instead of alleles of a single gene. You are also stopping your modelling at one generation and not factoring in all of the incoming mutations that occur while those mutations are moving towards fixation.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes: That's a computer simulation. How about an experimental example, one with real living things such as the Kishony or Lenski experiment but with sexual replicators? It's a computer simulation of a real population of fish, and the simulation matched the distribution of 3 alleles in 3 different genes associated with high salinity adaptations. Read the paper:
quote: HIV hasn't gone extinct and it doesn't go extinct in someone treated with 3 drug therapy. And your effective population size is only 20,000. I'm surprised you think that the real world isn't an extreme environment since 99% of all species have gone extinct. That doesn't sound like an evolution-friendly environment. More than 99% of bacteria die or fail to reproduce in concentrations of antibiotics above the minimal inhibitory concentration.
The reason why single drug-resistant alleles in HIV treatment don't increase in frequency is that these variants don't have reproductive fitness any better than the drug-sensitive alleles. If the drugs don't reduce the replication rate of HIV without the resistance mutation then what does the drug do and what is resistance?
Really? There haven't been famines, pandemics, droughts, wars,...? Which one of those wiped out more than 99% of humans? In what way were the post bottleneck environments stable enough to induce adaptation to them?
So 100% of the population all have the adaptive allele at one genetic locus and 100% of the population have the other adaptive allele at the second genetic locus. Why do they need recombination for a descendant to get both adaptive alleles. Everyone in the population already has both adaptive alleles. You tell me, you are the one who keeps talking about recombination events.
I've never written a paper on biological evolutionary competition other than to use Haldane's model along with my DNA adaptive evolutionary model to simulate and predict the Lenski experiment which it does very nicely. How does Haldane's model apply to different mutations on different genes in a diploid sexually replicating species?
This would be similar to what Haldane's Cost of Selection model addresses. His model gives a fixation rate for a single adaptive allele of 300 generations. Do 2 adaptive alleles at two different genetic loci fix faster than a single adaptive allele at one locus? Do 3 fix faster than 2, 4 fix faster than 3,..., 19,999,999 fix faster than 19,999,998, 20,000,000 fix faster than 19,999,999? I look forward to hearing your response to that question. They reach fixation independently of each other if they are not close to each other on the same chromosome, which is called linkage disequilibrium. If we take your 300 generation number, then in generation 1 there are 5 adaptation mutations and they reach fixation at generation 301. In generation 2 there are 5 beneficial mutations, and they reach fixation at generation 302. In generation 3 there are 5 beneficial mutations, and they reach fixation in generation 303. At no time does evolution just stop and wait for the previous beneficial mutations to reach fixation while ignoring all of the beneficial mutations that happen in the mean time.
How about a beneficial mutation occurring on someone from North Africa and a different beneficial mutation on someone from South Africa? That's only 4600 miles. Are there any long-distance relationships in your population of 20,000? It would depend on gene flow.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes: I thought that we were assuming that each member of the population were homogeneous at these two loci. Doesn't matter. For Aa bb x aa Bb, 25% of the children will be Aa Bb. That is true no matter how many other heterozygous or homozygous parents there are at the two genetic loci.
Do you want to show us how to compute the joint probability of one Aa parent mating with a second Bb parent to give an AB offspring from your population of 20,000 as a function of the distribution of different frequencies of variants? That is a multinomial distribution calculation with lots of different possible outcomes. Start with AA, BB, CC, Ab, AC,... You have a few permutations to compute. I already did that in a previous post.
Apparently, I've confused you. Adaptive allele A occurs at one genetic locus, adaptive allele B occurs at a different genetic locus, and the C alleles are the subset of all alleles that don't give an improvement in fitness at either genetic loci. Your math doesn't add up. Here is what you presented before:
quote: Non-disease alleles should serve this example well. There are disease alleles like achondroplasia (dwarfism), hemophilia, and cystic fibrosis. The non-disease alleles greatly outnumber the disease alleles. Let's say that each is at 0.99 frequency, and we will call them A, B, and C. So does your math make sense? 0.99A + 0.99B + 0.99C = 1 Does that math add up? Nope.
Taq, have you thought about what would happen in the fixation process if either the A adaptive allele or B adaptive allele gave greater reproductive fitness than the other? A would reach fixation in fewer generations than B. If fitness were additive then carriers of both A and B would be the fittest.
In the biological evolutionary competition, wouldn't the variant with the adaptive allele that gives greater reproductive fitness go to a frequency of 1 and the variant that gives a lower degree of improvement in reproductive fitness go to a frequency of 0? No. If that were the case then there would only be one non-disease allele and the rest of the disease alleles would drive towards fixation. You also ignore the fact, once again, that A and B can be put into the same genome in sexually reproducing species unlike what is seen in asexual organisms. You also apply the wrong formula which is meant for alleles of the same gene.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes: Let's say that every time you flap your arms, you generate a small amount of lift. I then take the laws of physics and formulate a computer simulation that determines the amount of lift as a function of the number of times you flap your arms. I plug in a number of times you flap your arms that is sufficient to generate a lift to get you off the ground. Does that mean you can actually fly? If the simulation is used on birds and it shows that they can fly, is that an accurate simulation?
But that leaves 1% still able to reproduce because they have the alleles that enable them to do so. Where in human evolution do you think there was a point where 99% of the population died off because they lacked a specific allele?
And in the case of HIV, this virus can do recombination. The problem for the HIV virus is that it cannot increase the frequency of any of the single drug-resistant variants to give a reasonable probability of an advantageous recombination event occurring. One parent is Aa bb and the other parent is aa Bb. 25% of their children are Aa Bb, carrying one copy of each beneficial dominant mutation. Do you describe this as recombination?
All these drugs inhibit the reproduction of the virus, they don't kill the virus. Recombination in the 2 drug environment might well get a two-drug resistant variant but the addition of a third drug (third selection condition) affects the mathematics of DNA (RNA in this case) evolution and random recombination for the virus to adapt. In the presence of the HIV drug, do the HIV resistant viruses reproduce better than the drug sensitive HIV viruses?
The point you are missing is that these bottlenecks select for variants with some degree of resistance to these selection conditions. For example, the black plague killed between 75-200 million people when the world population was about 450 million at that time. Which is all beside the point. All I said is that bottlenecks aren't necessary, not that they never happen.
You can't have different mutations fixing simultaneously unless you have hitchhikers. Haldane's math applies to the fixation of a single allele. You want to consider two different adaptive alleles at two different genetic loci fixing simultaneously. But that only allows for a frequency of 0.5 for each variant. That is only for alleles at the same genetic locus, not for mutations at different genetic loci.
So the frequency of the A variant is 1 in the population and the frequency of the B variant is 1? You need to check your math. Are you saying there is no base in the human genome where nearly 100% of humans have the same base? What are the frequencies of the wild type allele for genes where there are known Mendelian diseases like achondroplasia or hemophilia?
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes:
Any time you are ready to get beyond Mendelian Genetics, let us know.
When you are ready to stop rejecting Mendelian genetics, let me know.
Are we to understand that in your population of 20,000, the only possible inheritance in that population is Aa bb x aa Bb? aa bb x aa bb can't occur? Already explained it in the other post.
You should know better than this Taq. You can only add frequencies (probabilities) if they are mutually exclusive. Then you should know better, too. So why are you using this equation for mutations in different genes when it clearly does not apply?
The A variant would drive the B variant to extinction unless there happened to be a B mutation hitchhiking along with an A variant but that AB variant would have better reproductive fitness than either the A or B variants. Are you the same person who wrote this? "You should know better than this Taq. You can only add frequencies (probabilities) if they are mutually exclusive." A variant and B variant are not mutually exclusive in the same way that the non-disease alleles are not mutually exclusive.
I don't ignore the fact that A and B adaptive alleles can be inherited by an offspring in a sexually reproducing population. I'm saying that the probability of that happening depends on the frequency of those individual variants in the population. The strange claim that you are making is that both the A and B variants can go to fixation simultaneously. What is so strange about that if the mutations are in different genes? You already accept the non-disease alleles all being near fixation, so why not mutations A and B found in different genes?
This is a point you still have to learn about mathematical modeling and computer simulations. You have to test your model against experimental data to test the veracity of your model or simulation.
The computer simulation has been tested against experimental data:
MetaPopGen 2.0: A multilocus genetic simulator to model populations of large size The human population size is too small for any significant DNA adaptive evolution, bottlenecks or no bottlenecks. Assertion without evidence.
Are you now claiming that a parent only passes a single genetic locus, not an entire genome? That's what you are claiming, not me. You claim that the frequency of any two mutations anywhere in the genome can not add up to more than 1. Obviously, that is false.
The numbers I've heard are that humans have in common over 99% of our DNA. But if you are going to add frequencies, make sure they are mutually exclusive.
A million irony meters just exploded.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes: When did I reject Mendelian genetics? You reject Mendelian genetics when you require the percentage distribution of alleles in different genes to add up to 1. You keep doing this. I bet you do it later in this post.
Why can't you add these frequencies? You have 3 subsets in the population. One subset has allele A at one genetic locus. Another subset has allele B at a different genetic locus and a third subset has neither allele. Unless there is an intersection of any of these three subsets, they are mutually exclusive. Each of these 3 subsets has a subpopulation size associated with their corresponding subset and if you add up these subpopulation sizes, you get the total population size. And there it is. You still don't understand basic Mendelian genetics, at least at the time you wrote this post. Let's see how this works out. Let's use 3 hypothetical disease alleles (e.g. cystic fibrosis, achondroplasia). Let's say 1 out of 1,000 people carries the diseases allele, or 0.1% of the population. This would mean that 99.9% of people don't carry the disease allele but the healthy allele. Let's call these Aa, Bb, and Cc alleles for genes A, B, C. According to you, I should be able to add up the number of people with the A, B, and C allele and get an accurate population count. Is that true? NO!!! If I do as you suggest I will get about 3x the actual population number. Do you know why?
Fixation by selection involves the most fit allele going to a frequency of 1. Other adaptive alleles that give a smaller increase in fitness are not increasing in frequency. You still don't understand how sexual reproduction works.
How about every real, measurable, and repeatable empirical example of DNA evolution? Start with the Kishony and Lenski experiments, then go on with the success of combination herbicides, pesticides, and rodenticides inhibiting the evolution of selection pressure resistant variants. How about mathematical models that predict and simulate the behaviors of these evolutionary processes? How about learning how genetics works? Why don't you start there? You seem to think that there is just one gene per genome. Perhaps you could start by learning how meiosis works, or perhaps what a chromosome is. Maybe you could learn that there are multiple genes in a genome, each with it's own set of alleles.
Me: You claim that the frequency of any two mutations anywhere in the genome can not add up to more than 1. Obviously, that is false. You: Don't be silly, I haven't made a claim like that. And of course, frequencies are always less than or equal to 1. Frequencies range from 0 to 1. Why do believe that there can be frequencies greater than 1?
So you claim you don't make that claim, and then make that very claim in the next sentence. You effing moron. Edited by Taq, .
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Quiz for Kleinman:
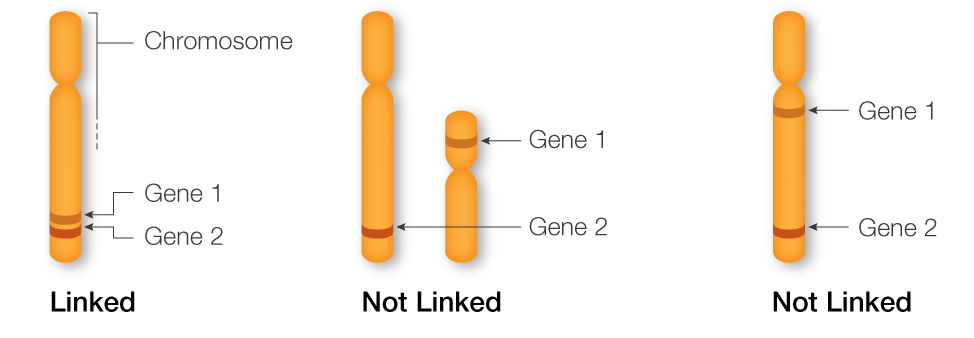
Why are gene 1 and 2 linked in the first example but not in the other two examples? What effects does linkage have on relative allele distributions between linked genes compared to unlinked genes?
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Dredge writes: If you flip a coin three times, for example, each flip is an indepedent event. How would you calculate the probability of getting 3 heads from 3 flips? We are getting more distant from the actual genetic system that got this all going. Kleinman is claiming that if we add up all of the allele frequencies for all genes in a genome that we will get 1. That's ridiculous. That shows a massive misunderstanding of the basic mathematics of genetics. Let's just say that there are two alleles for each of the ~30,000 genes in the human genome, and the frequency of each allele is 0.5, or 1.0 for each gene. If we add them up like Kleinman claims we can then we get 30,000, not 1.
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1
|
Kleinman writes:
Don't hold your breath for Tany's explanation of how DNA evolution for viruses, bacteria, and yeasts differs from DNA evolution for complex, multicellular, sexually reproducing organisms.
We could start here:
|
||||||||||||||||||||||||||||||||||||||||
|
Taq Member Posts: 10085 Joined: Member Rating: 5.1 |
Kleinman writes:
This does occur but you have to have very large populations in that family. That's why universal common descent is not possible for primate precursor/human-chimp evolution. There isn't a large enough population size for adaptive evolution to operate to account for the fitness differences between humans and chimps. Taq understood this and tried to address this problem with recombination. But random recombination is also governed by the multiplication rule. Taq tried to address this with a physically impossible claim and he knew it. That's why he abandoned the discussion.
Bullshit, K-man. Life happens, and I got busy elsewhere. Let's revisit this classic you used in post 311
quote: Do you still stand by this? Do you add the allele frequencies for alleles in different genes? What about the healthy alleles for 3 different genes associated with a genetic disease, each with a frequency of 0.999? 0.999A + 0.999B + 0.999C != 1 OOOOOPPPPPSSS!!!! Your math doesn't work.
|
||||||||||||||||||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024
 (1)
(1)