
|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| Thread ▼ Details |
|
Thread Info
|
|
|
| Author | Topic: Are human tails an example of macroevolution? | ||||||||||||||||||||
|
Brad McFall Member (Idle past 5063 days)  Posts: 3428 From: Ithaca,NY, USA Joined: |
I would be contending that the "phylotypic stage" does not homogenously exist across the psm. I would revert to the Elder Darwinian, Erasmus instead who used the word PLAN(in the section quoted in Carter I displayed interthreadalia), which now would mean(for me at least) the split(left and right)-somite-formation, whether clocked or not, rather than the current version that the adjective("polytypic") derives as to a "body" plan which is Logically referable to Woodger's use on any dissection I should think( unless I am mistaken in this attribution). I do not see how short of prejudice one is out of the woods on bilaterally symmetric animals here, SINCE the somites themselves divide among left&right.
I can see how a reductionist would try to read the parallel signaling paths as a means to remove this ambiguity but that does not seem to mean that it MUST be extended to the "tail bud." There just is not as much information on the tail bud as there seems to be speculation about recruitment INTO the PSM. I say all of that without thinking about cell death in this context. You said, quote:but even if the VER looked like a limb bud, where is the evidence sans speculation and hope for future research information that, "it appears" that Hoxb-13b stops tail development when the creature has a bud but nothing tail wiser. In other words how can be certain that if Hoxb-13 does not stop "overgrowth" (even though it appears to in tailed creatures) that it is not neutral or functions differently in a differently familiar situation where the development does not normally yield a "tail"? I have not looked THIS up, but on reading that the PSM can be inverted without directional periodicity being upset could not the Hoxb13 effect affect at this "stage" rather than the difference of anterior and posterior PSM which have different signaling relations anyway?? You seem to supporting the contention that the tail embryogeny must be historical by anatomical divisions which I do not see supported by data as it exists. I do not deny that the literature seems to be written with this idea, as a goal or target but I can sense how the information portrayed in the actual literature is nothing other than a feeling for belief in evolution rather than discrimination within evolutionary thought that challenges if the data really supports an historical contingency rather than a formal configuration both of which may or may not remain purely "evolutionary" when retained in sustained thought. I am happy to see that readers of EvC might be beginning to notice a tangible difference between yours and my views and do not be discouraged or fell timid about asking to me answer other questions you have already asked. We have covered a lot a ground and I am more than happy to revisit older posts and other places I said I would update. I just do not have a lot of time as we all do not so I felt posting something was better than little to an avalanche later style. You are correct I misread "d" and "b", my bad. That post however was a general romp though "hoxology" but I am going to try to let Gould go if I consciously can and just talk to you from me. I will refer back to his schism between positivity and negativity if I need it but as you have done a great job pointing me to sufficient sources to carry out this dialogue I pray they were so indeed. I do see that there is some theoretical "tension" between multiple signaling pathways in the caudal end and the space and time correlations FROM homeodomains so perhaps I am premature here@Best, Brad. Edited by Brad McFall, : spelling
|
||||||||||||||||||||
|
Brad McFall Member (Idle past 5063 days) Posts: 3428 From: Ithaca,NY, USA Joined: |

quote: The idea (“core effector mechanism”) upstream and the view that genetic programs allowing regulated cell suicides in single-celled organisms (also discussed by Amesien) extends the whole matrix of the “quirky functional shift” (when applied to an ”organ’ rather than a ”social organism body’) into a nexus of thought where it is far from clear whether one is able to get on the side of the altruist or selfishist. Thus when I challenge that the embyrogenic facts bear more on recruitment OF cell lineages INTO the area of the PSM than of the tail bud lineages cellwise one has a hard time sustaining a thought linked to Amesien’s dissection of various levels (cell vs individual organism) in an evolutionary perspective of cell death no matter how the bacteria competed or not. Thus without that, it seems to me that cell death in whatever way it might be retained genetically could be providing simply a space or rather lack of cell area to morphogenetics rather than being a positive construcutor IN THE TAIL (the end) even though this would not necessarily simply be such a bound and is rather constitutive for other areas of a developing body. I have read somewhat of the cell death in the tails of frog tadpoles and in reading that literature I have only been lead to think that cell death “destroys” a morphology in order to provide another (stronger back legs than front legs (to the gills)) (altruistically) and not that it regulates a part (tail bud) for the function of the tail itself(which seems to be necessary if the human tail is to be explained by the same history that gave up the frog’s lack). This thought gives a purely structuralist explanation for cell death in the tail of a vertebrate and removes even the functionalist one. Here is how I thought of the idea of cell death in plants. It could be related to that in animals via the right angles in the plant if the nodes intercalate(interfinger) the spatialization of molecular reaction pathways in the animals. I have added a page TheTrainer's pregram which contains comments that fully spatialize cell death as I understand it and explains a bit why this idea has not been developed as of yet. If hoxologos is the correct arrow for this quiver in this particular threads case, I will have to adjust that page at least. Edited by Brad McFall, : subtitle changed for information Edited by Brad McFall, : plant idea added Edited by Brad McFall, : with link to external ideas on how cell death relates to any soma
|
||||||||||||||||||||
|
Brad McFall Member (Idle past 5063 days) Posts: 3428 From: Ithaca,NY, USA Joined: |
There are some difficulties of language going on here. When I quoted Carter on Darwin I had in mind his anglo-expressions. I had hoped that by moving THROUGH Gould's lingo we would have exhausted our likely different linguisitic backgrounds. That does not seem to be the case given you response.
I was reading "stage" in an general evolutionary sense. The general point is that I can not see how the topography of embyryogeny presents a topology that *must* be historical. Your reply in this post NOW is about me simply reading text(s) you provided. The questions are not of the same quality as before. The internet might have others thinking differently and mistakenly thus. Yes tail somites ARE formed from the psm, as I read, but the "bud" exists as labeled beyond and BEFORE the somite pair appears as it appeared in the figure one, I think. Am I seeing things wrong? Are not the two "rods" of tissue not each on a side off the a drawable midline?? In frogs the macophages also destroy the tail but I do not see how the tail exisiting outside the psm in humans indicates a "lower" stage evolutionarily, necessarily. The tail is simply tissue at the end of the creature. If there was some archetypical difference between radially symmetric and bilateral organisms as some creationists may wish to claim, then there might be reason to argue immediately from the last place in the tissue as due to constituitive form-making. The psm marks the place between this edge and "interior" tissue. As for what "may" (MUST(sic!)) be extended, they are(were) the signalling pathways however interpreted. If you can show me that hox molecular biology(relations of hox cluster physcial chemistry as promixate cause of all allomteric regularities) regulatory for the tail bud tissue itself then yes, I might be inclined to accept an historical interpreation for the functioning of the this "organ" or "tissue area" and would. Perhaps I have misread the hypotheticals with regard to the pathways but If one intends on also involving the evolutionary notion of cell death rather than a simple list of genes involved in cell death itself it seems that anatomy will be even more difficultly supporting the historical falsification of my view or one similiar. It was not necessary therefore that I thought the somites "divided" left and right but only that there was (INTERNAL TISSUE) between the period when a new somite "pair" develops even if there were some asyhchornicity. The issue of how the EDGE gets divided from the zygote is a real one. I notice the topological issue is probably at the blastula to gastrula stages, saying it plurivocally. Maybe I missed it, I am not trying to be obtuse, but I do not see how the existence of HOX cluster implies that the the terminal segement area must be non-scalar with respect to the magnitdue of a matrix metric that links(Correlates) the parts UP TO the END (of the creature). This would not apply to the internal structures logically however. Sorry about msg 76 I will back over to that if I knew which thread it was in?? this number is 54 I take it your position is that if it was not for cell deaths which destroy the end/free embryonic tail no matter how much was overgrown by the trunk that, humans would posses in sexual maturity, a tail, as sometimes has been clinically recorded in a some hundreds at most individuals and because there are some deeply conserved issues of segementation between the vertebrates and invertebrates the end member of a serial division of the anatomy from head to end necesarily implies that the region between the free end the "trunk" must be explained as simply a matter of historical continuity from a heritable program saved over the generations? Do notice that there is "gut" tissue in this end region. When thinking about tadpole tails I was able to think about ecological uses of the tail itself. Are we certain that there is not some gestational use for a free tail in fetuses? Could the few cases of adult human tails not be due rather than one associated with hoxology to one of overexpresion of genes possibly associated with a behavorial cause of embryonic free tails (assistance in untangling the umbilical?)? 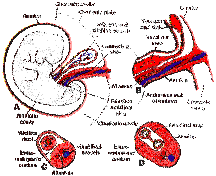
quote: Edited by Brad McFall, : msg number issue Edited by Brad McFall, : small words Edited by Brad McFall, : generalization Edited by Brad McFall, : umbilical link
|
||||||||||||||||||||
|
Brad McFall Member (Idle past 5063 days) Posts: 3428 From: Ithaca,NY, USA Joined: |
According to the E. Darwin info I used in message 9
EvC Forum: Are human tails an example of macroevolution? 1st- metamorphosis 2nd- artifical selection or cultivation (in the garden)(Mendel distinguished this from cultivation in the country) 3rd - meteorologically torque affected biochange or nonadaptive trait procurement - habit via employment (Marxist) - hybrids and or subject of this thread 4th - WHEN WE OBSERVE(D) the essential unity of all plan of all warm-blooded animals WHAT THE FILAMENT WAS as believed. The FOUR that Bernd
http://EvC Forum: Are human tails an example of macroevolution? -->EvC Forum: Are human tails an example of macroevolution?
was want to express from E. Darwin was quote:already had the triple 3rd above, which I asserted only makes sense ON ROTATION. Bernd is challenging that my cross generational abduction (between points two(plants) and three(topic at hand)) is never physically rotated in the revolved thought of any stripe down the right and left at an terminal ordination of the datum. I think it is. There are horrendous difficulties in communicating in this thread. He now said that his own thoughts did not matter. He contends that this is a ready read thread or filament of ANATOMY not of phylogeny. I can only see this as possible if the connections of nonadaptive traits completed by seasonal and climate effects only dominate the "similarity of forms" and thus I find that this is actually an issue of translation between languages rather than a issue where one is trying to decide if the evidence pertains to a structural or an historical angle (or functional) in explanation of the clinical cases. He now redirects back to my own ideas. This is always easier to answer than to argue in general. I will. But you can see now as this thread has come and gone and goes some more that it is definitely NOT easier to sustain a creationist position vis a via a particular topic on EvC!! The forum is with C. Darwin's not E. Darwin's view. Is the prosimian tail the Homo tail phylogenetically? C.Darwin indubitably took "similarity of form" as Bernd shortened it quote:http://EvC Forum: Are human tails an example of macroevolution? -->EvC Forum: Are human tails an example of macroevolution? and added the notion of "history" precisely as Gould has narrated this past science somewhat but if one reads very very closely Charles Darwin's Power of Motion in Plants quote one can read that orthogonal directions decompose OUT the form of the seed. So if Mendelism changes C.Darwinism Within the third place (INTO the second) the "structure" that is the difference of the numerator and denominator can "constrain" the "Form" in the induction beyond E. Darwin IF the tail is read as the filament or flaccid rope (herein and of Galelio etc). Bernd continues to suspect that I am reading something into this that is not here. I will show him I am not. Bernd already agreed that he will take into consideration "heritability" on the "family" level. Making this case out clearly is beyond the pale of what goes down at secular elite universities.
|
||||||||||||||||||||
|
Brad McFall Member (Idle past 5063 days) Posts: 3428 From: Ithaca,NY, USA Joined: |
Perhaps I was mislead a bit by the labeled picture of TBM in a link upthread.
I do not "visualize" evolutionarily that there was a larger "anatomical diversity" of life prior to the Permian as did Gould. Instead when we have to dicuss all of the creatures I have HAD this of "gestalt" in mind.  I probably constructed this template during my earlier years posting on EvC. It was probably NO COINCIDENcE that I cut out the tail end of the frog area TO BE ABLE TO ACTUALLY FIT IN the word "data." Now if you feel I have still overridden the evidence from hoxology please let me know but under a term "transmorgification" I would still retain that cell death can possibly be theoretically capable of giving PHENOTYPES space. THE ACTUAL SPACE was triplicated by the brown /\'s in the following (notes on the relation between any given angle, behavior and canalization. I had thought that cell death is just a bunch of those symbols signified (but "programmed"). It may be possible that negative entropy is involved but after communicating with Gladyshev I am less prone to go there (source vs sink) as easily as I was when I first scrathed down the relations. This was why I was saying that it might not matter to you but it does to me if the canalized pathway is wide or narrow. Obviously these are only my own marks. By 'Stronger' legs I simply mean that frogs have back legs to jump with and the anatomy is proximate to the tail rather than the gills as to where the macrophages and destruction is going on. I am not sure that "tissue remodeling" and "transmorgification" (as intended by Gould) are any different. I did find some of the herp papers on somites in lizards I had mentioned earlier. You may note as to your other reply to me that I mention "vectorization" in the white picture above. I also say that this view of mine has not existed. Part of the interest in this relation of behavior to morphogenetics (across a mathmeatical construct of my own doing) was discussed with BEN (in part at least) when he first started posting on EVC. I can admit to this much.
|
||||||||||||||||||||
|
Brad McFall Member (Idle past 5063 days) Posts: 3428 From: Ithaca,NY, USA Joined: |
The idea I got on reading some of the material you linked was:
There is always some part at the terminal "end" of the vertebrate embryo. I was trying to generalize beyond the vertebrates if and when I mention Agassiz. But so that we can maximally understand each other lets drop retrodictions to the very beggining of this thread unless it is the last position of discussion. I will assume we are only talking about vertebrates (not even echinoderms) and never about nodal space differences in plants that twist. In the picture I was looking at there were no somites formed yet into the psm and the tail bud was labeled. I noticed that the the somites were also marked with with dots on each side of the axis of the back midline. In what I had thought was my last post in this series I suggested that BEHAVIORAL functionality may ADDITIONALLY format the embyronic functionality of this area and that instead of (as well as in addition to ) hoxology being the doxology of human "tails" rather there may be some deviant gene overexpression for this 'behavior' that extended beyond cell death protocols that would normally prevent the trait from appearing at birth. Now there may be molecular evidence contrary to my own analysis and somewhat lame synthesis. But this property of the free tail may indicate a structural origin of the LACK of the great ape tail even while it faimiliar structure for the monkeys and other mammals as a whole. In this case I would have taken the destructive notion beyond the material destruction (as I have thought with frogs) and implicated gestational behavior as controlling genetic fate as well. Look I dont deny that the tail might be historical as Gould is want to disclaim against Dobshanky's "topography" of lions and tigers, and wolves and jackals but I only wanted to make sure that there was not some other possibly wholy structural explanation that just might not have been recieved into the current literature. I do not really feel we have made it to the place in the discussion to cause me to get even more rigorous with the "linear transformation" etc but I can go there if that is really what you are calling for.I thought the reference to "parallels" would suffice still for now. I, for my self, have a hard time trying to disuade myself that you are saying nothing more ( with great links to modern literature notwithstanding)than:
quote:but look I have not tried to assimilate ALL of the hox literature. I am reading Gould rather deeply at this time and my confidence comes from mental sparring with his ideas on the subject more than what others may feel is possibly more representative of the current literature. I am not making apologies. I may be wrong.
|
||||||||||||||||||||
|
Brad McFall Member (Idle past 5063 days) Posts: 3428 From: Ithaca,NY, USA Joined: |
Dear Bernd;
Your reply seems well (enough) balanced. Yes, in a sense "the weight" of the literature slants in your direction. The best I can do is present what goes on in the(an) intuition that bends another way. If I was wedded to my own thought process I would be challenging just how much we know about the"spontaneous movements"(lack thereof at a time needed) you indicated as falsifying my own conclusion. I rested with that post you quoted from because I was merely trying to point out an alternative way to think about the subject. I think you have seemed to have taken into account the heritability or potential of such at the family level (but you have not addressed this specifically). I think it is not an issue that if one was to objectively compare my own way of thinking biologically and some other way that is already instutionalized, I would loose, in the comparision. That is always a given however. So, INDEED, if my argument hinges on the "behavioral" movement of the embryo (and it was true that there is no actual movement as you assert) then yes it would not be unfair to say that there is more "supporting" evidence on your side. I will go back and try to read hoxology more critically because you seem destined to recall the conservation of hox genes in what I would otherwise simply think is the "topology" of tissue formation. Inother words, I will not consider this thread finished and I will try to show you once again that a structrual possibilty is not off the table. We will have to visualize cell divisions in this case. There is not a need to worry about the temporality of your postings. I am reasonable. I have been unable to find the post on EVC where I first started to discuss the topology of cell divisions from the zygote. What is important is to "see" how space(inside vs outside) becomes "a scarce commodity" after the first dozen or so divisions before and beyond blastulation. My intent will be to show that tail space is ALWAYS "outside" in a topology so describable and is thus in a different logical position than internal tissues when discussing the data of evo-devo (it is not simply another "section" as concieved from insect anantomy but is "terminal" and thus need not be subject to the same spatio-temporal chemical affects as can be resonable effected on the aggregate). In other words I do not take the vertebrate seriality and invertebrate sections as necessarily coincident as geometry would trend one to think from a common mouth. I will suspect that aspect of any instutionality of hoxology. Insects look more like hybrid plants than lethal genes to me. But that is just me. What is required is to seperate Bauplane and Plan. I feel that current elite biology on either Mayr or Gould's bird watch fails to distinguish trascendental reflexions of form from any meaning of plan readable in Carter here no matter the watchful thought of OT Isaiah. Notice that Carter speaks for a large history of German thought. Without intricate trichotomies of creationist thought the words "bau"PLANe and plan simply 'run together.' I think that the only way to draw the corrrect lingo would be to work with Woodger's "biology and language" of Russel (functors) but we will not be able to do this in this thread. Instead, we will get the chance to expand on interpreting evo-devo. I suggest that Gould's reading will not distiguish the reading from German "Natur(-E)-Philosophy" in the plan the sari of this thread woofs. The sentence I find conflated by the best of the best in all possible worlds of evolutionary thought was:
quote:
quote: Edited by Brad McFall, : No reason given.
|
||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024




