|

Understanding through Discussion |
|
|
Register | Sign In |
|
QuickSearch
| Thread ▼ Details |
|
Member (Idle past 1435 days)  Posts: 20714 From: the other end of the sidewalk Joined: |
|
Thread Info
|
|
|
| Author | Topic: Interweaving Evolution & Hybrid Vigor | |||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
I was rereading the Peter & Rosemary Grant, Darwin's Finches and Evolution thread and in particular noted this passage from the initial quote (Message 1):
quote: And I also recall the discussion on New Species of Homo Discovered: Homo nalediHomo[/i] Discovered: Homo naledi of a "braided" history ... from the article:
quote: Because Homo naledi is a mosaic of features some modern derived features and some preserved ancestral features, and that applies to other species, such that there is some mixing and matching going on, this suggests some hybridization in the past. We also know from DNA analysis that there was some hybridization with Homo neanderthalus (alt Homo sapiens neanderthalus ) Wikipedia defines Heterosis as:
quote: An example of outbreeding depression would be horses, donkeys and mules, where reproductive isolation is not complete but usually results in sterility. So outbreeding can result in hybrids that are more fit, equally fit or less fit, at which point natural selection would favor the more fit (heterotic) varieties becoming dominant in the population/s, leading to a new species group. Crossbreeding is done a lot in animal husbandry and agriculture to develop improved stock, so it should not come as a surprise that it can occur naturally when daughter populations regain contact after undergoing different evolutionary experiences, but before they have evolved reproductive isolation. The amount of independent evolution could vary a lot, as it is the development of reproductive isolation that would stop such interbreeding. Certainly when we see that hybrids can be made between lions and tiger or between lamas and camels after long periods of separation. We also see this between European invaders and Native peoples of the Americas. So the tree became a bush becomes an interlinked\braided bush. A Where C is not the same as A, but is a braided mosaic of B and D. Note that A, B, C and D still form a clade descended from A. This does not mean that evolution does not happen, just that the process is not a cut-and-dried cookie-cutter proposition. This also means that the definition of "species" is a little muddier than before ... and it was muddied before. Enjoy Edited by RAZD, : subt Edited by RAZD, : .. Edited by RAZD, : fix code Edited by RAZD, : .by our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Another application of this to the debate is when we look at animal husbandry, agriculture and breeding programs.
So outbreeding can result in hybrids that are more fit, equally fit or less fit, at which point natural selection would favor the more fit (heterotic) varieties becoming dominant in the population/s, leading to a new species group. Now we can see in dog breeding that some dogs are bred to create an extremely inbred population in order to maintain the "breed" and that these types of breeds often exhibit inbreeding depression. These are the "show" dogs, but they are not the only breeds. Another class of dog breeds are "working" dogs, where the appearance of the dog is not as important as the function. The sheep herding dogs come to mind. To my view they seem to exhibit Heterosis and not exhibit any of the inbreeding symptoms. We can also look at cow and pig breeding, where the purpose is not some aesthetically "pretty" animal but one that fills a function (more milk, more meat). From the Wikipedia definition of Heterosis again:
quote: When the aspects of rapid growth, size and robust health are considered it is easy to see why this is pursued in husbandry. Again, we can see how (properly applied) artificial husbandry practices offer a window into how natural evolution (mutation and selection) can operate. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Punctuated equilibrium - Wikipedia
quote: The standard view iirc, is that a small subpopulation evolves in isolation, and then returns to the habitat of the parent population and displaces it with their better adapted phenotypes. With the emerging thoughts on "braided" beginnings to new species and hybrid interactions during the early stages of new species formation, it seems more logical to me that the returning subpopulation interbreeds with the parent population and the hybrid offspring inherit traits from both populations, which are then selected for fitness, leaving a new population with hybrid vigor and a mosaic of traits. This has the benefit of mixing the best of both populations within a larger breeding population. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Discussion, on how evolution works in regards to speciation. It's not always straight line process of speciation, but a little tangled, a little messy.
Also, I think punctuated equilibria when a population experiences a period of rapid evolution, as opposed to relative stasis. The population could subsequently return to displace the original parent population, but I think that's just competition, not punctuated equilibrium. The basic idea behind punctuated equilibria, as I understand it, is that a smaller relatively isolated population undergoes rapid evolution, possibly to adapt it to a slightly different ecology. Then it returns to the parent ecology which has been in stasis (continually readapting to a static ecology) and has an advantage for survival or breeding, and then it takes over and displaces the parent population. The question I am raising on this is whether full speciation of this daughter population is necessary, or would not a new varietal phenotype that can interbreed and form hybrids that join the best features in a mosaic of phenotypic traits be a valid or better explanation. We have evidence of such mosaic evolution, the braided stream pattern noted on the Homo naledi thread for instance, and the Darwin Finch hybrid "big Bird" as another. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Possibly. Or it might migrate somewhere else. Or nowhere else. Or it might return to the parent ecology and co-exist with the parent. Or it might return to the parent ecology and find that there is no longer any parent population to compete against because it's gone extinct. True. Part of the reason I want to open this to debate with others, particularly those that know more about this than I do.
I thought what was novel about the punctuated equilibria idea was that the episodic nature of the fossil record wasn't necessarily just an artifact of an incomplete fossil record, that it could also be real. Indeed, but for my argument ("the part I want to focus on" ... ), especially in those areas where successive fossil layers show noticeably different forms. Now, I would like to credit Faith for leading me to think in this direction while formulating responses to her argument and her questions on this phenomenon in the fossil record. And I have another example that I am working on, which I will save till after promotion and some confirmation from others that I am not just blowing smoke. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
In Why Evolution is True, There are no ring species Jerry Coyne argues that there are no "ring species in the classic sense" ... and then goes on to describe what he feels the "classic sense" is:
quote: The idea is that classic ring species demonstrate evolution in a horizontal space continuum as opposed to "traditional" evolution in a vertical time continuum, and thus we can look at the intermediate stages between initial division of a population until fully independent species are developed, which in the classic ring species would mean that the two end points don't interbreed. He further describes this classic ring species as a continuous gradualistic development:
quote: He then cites why the classic ring species does not exist:
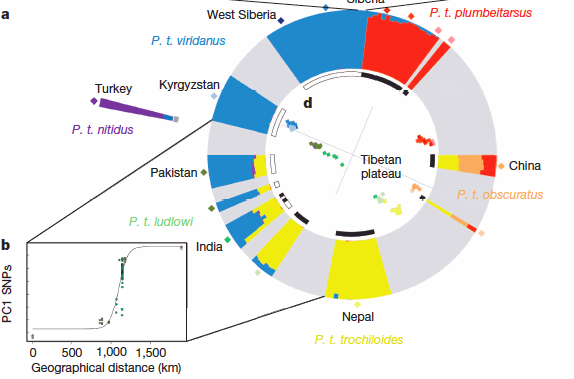
quote: So not a classic continuous gene flow ring species ... and all these examples show lumpy evolution with some isolation and rejoining of the ring variety populations. Messy. Here's another graphic from the Genomic divergence in a ring species complex paper that Jerry cites:
When I look at these diagrams I see four genetic groups, P.t.nitidus (purple, the western-most, isolated group that may have been the parent population), P.t.viridanus (blue, perhaps the founder of the western clade), P.t.trochiloides (yellow, the first described "type" of P.trochiloides?) and P.t.plumbeitarsus (red, perhaps the founder of the eastern clade) that are pretty much homogenous, and that P.t.ludlowi appears to be a hybrid between P.t.viridanus and P.t.trochiloides that has evolved some distinctive traits (green patches) not shared with either parent population, and likewise that P.t.obscuratus appears to be a hybrid between P.t.trochiloides and P.t.plumbeitarsus that has also evolved some distinctive traits (orange patches) not shared with either parent population. It also looks like a hybrid population may emerge in the north where the ring ends meet and some interbreeding occurs ... or the eastern variety will increasingly become a hybrid with the introgression of the western variety and the loss of gene flow from the south due to habitat destruction. Only time will tell. Jerry concludes:
quote: Indeed, "species" is a matter of definition, and if we apply the definition of species discussed on How do you tell one species of turtle from another? where the two populations show distinct genomes and interbreeding is not considered a deal killing element, then it looks to me that we have four species:
... if we use the "genetically distinct" definition for species. How does this fit in with the "Interweaving Evolution and Hybrid Vigor" argument? Well, we now have two large populations of hybridization that have evolved some distinct traits not found in the neighboring populations:
... with further small hybrid zones between them and their neighboring populations through geography that tends to isolate these two large hybrid populations. The small zones appear have waxed and waned, but they also seem to allow some gene flow when they active, so the two large hybid populations can have limited interactions with the neighboring genetically distinct populations. These two populations show exactly the intermingling and then later differentiation evolution posited in Message 1.
So the tree became a bush becomes an interlinked\braided bush. A Where C is not the same as A, but is a braided mosaic of B and D. Note that A, B, C and D still form a clade descended from A. So, not a "classic ring species" but an example of horizontal braided mosaic mixing before species are reproductively isolated. Enjoy Edited by RAZD, : clrtyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
I wish there was a way I could get my argument onto this thread since it is touching on so many areas of interest to my point of view, but I can't think of a way as it is currently written. Well I would like to finish making sure you understand that I understand your argument by finishing the discussion on A New Run at the End of Evolution by Genetic Processes Argument (see Message 209 latest post). Then we can see how it applies or doesn't apply here. Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
I think you may have a slight misconception about punctuated equilibrium. The basic idea behind PE is that organisms remain relatively stable for long periods of time and then make a sudden evolutionary leap. ... Where that sudden leap occurs in a small population, partially to totally isolated from the main static population -- that small(er) population can evolve faster due to the smaller gene pool where an advantageous mutation\adaptation to a secondary ecology can spread relatively quickly.
... and then disappearing from the fossil record to be replaced by a derived version. ... Yes, that came in from the periphery ... ... but was the old population replaced or is it a hybrid population, mixing the best adaptations from the isolated population and the static population? Certainly the derived new population is a mosaic evolution from the static parent population with some new traits and some old traits,
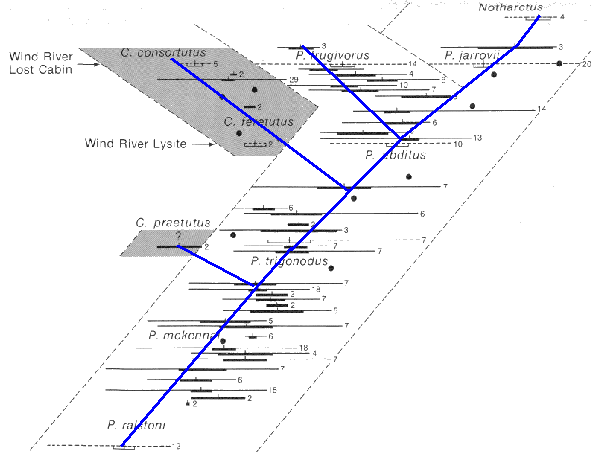
... It is often thought of as being in opposition to gradualism, but in reality it is a form of gradualism. Or we see a range of evolutionary rates and selection pressures. Certainly when we look at foraminifera the record is one of classic gradualism occurring over generations, and certainly when we look at pelycodus we see a general gradualistic overall trend from small to larger, and then at the point of speciation there is a 'sudden' (eg quicker) trend back to small in the one branch to provide 'distance' from the other branch (and resulting reduction of competition) ... ... and there were other earlier branches that failed and either died out or were reabsorbed into the main population. Doesn't that have the potential to introduce new traits back into the main population? For instance here are a pair of different interpretations of pelycodus and copelemur evolution:
In both interpretations the first branch appears to die out or get reabsorbed, and I (not surprisingly) favor the reabsorbed with new mutations added back into the main population. Also I see that the whole population shifted to the left as this occurred, which is another reason why I see reabsorbtion as a viable option here. Side note: Copelemur is named for Edward Drinker Cope of COPE's Rule that species over time will tend to increase in size, a trend we definitely see running up the right side in both images.
Travelling towards the center of the sphere is adaptation toward an optima. The red line is the fitness path a hypothetical organism might follow. Sometimes the path doesn't lead directly to a most optimal level, but makes a big change not directly towards the center. Some changes are quite small, but others (one in particular) are rather large. The large step (the third layer in from the outside) is what Gould would have identified as punctuated evolution. Another observation Fisher/Orr made was that as the fitness gets closer to optimum, the steps begin to get smaller. So, there is not likely to be large leaps when the population is close to optimal fitness. I think Orr has a good balance between PE and pure gradualism (including neutral theory). As long as the ecology stays static the selection pressures will be static, and large populations will tend to select towards the centers of their ranges of variations -- ie for stasis. The evolution of variations will be more tolerated at the edges where less viable individuals are pushed into marginal habitats, and if the population is large enough it will cover multiple habitat\ecologies and you can get population subdivision with anomalously high proportions of homozygotes compared to completely homogeneous populations. or one could say ... evolution happens.
The phenomenon you are describing is called reinforcement.
This appears to be an excerpt from Jerry Coyne's book "Why Evolution is True" and is a good article about reinforcement. Super nice links, I spent a couple hours perusing the first back to the A-Z links and all the items listed. Sad to see it did not list 'mosaic' but not surprised. One of my dad's pet comments was that evolution in general and human evolution in particular displayed mosaic evolution, essentially where parts evolve rather than the whole individual. He thought it was underplayed in the field. But no, not reinforcement -- as that leads to speciation, but rather an interweave where different populations rejoin and form viable hybrids that exhibit hybrid vigor and how does that occur in natural systems\selection. Consider the level where populations rejoin, there are genetic differences due to isolated mutation selections, but now they are interbreeding -- cross breeding -- and re-fertilizing the static parent population with new fitness traits that take over. This introduces new traits into the population with a wave effect. Or am I smoking some kind of crack? Enjoy Edited by RAZD, : addedby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
Again thanks for the links, the Evolution - A-Z link in particular is an excellent resource and should probably be posted in Links and Information.
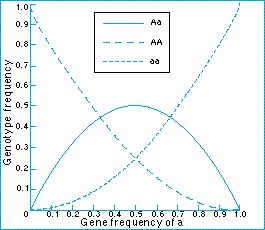
The Hardy-Weinberg ratio explains quite easily why mutations or rare alleles are more likely to be spread in a small population than a large one:
quote: Now I was a little confused at first by the "reached after a single generation" comment, until I realized that this applies to each generation and changes with selection acting on the relative fitness of p and q. This also assumes equality between male and female viability and distributions, as the probability of the offspring combinations is the probability of male (p,q) mating with female (p,q) and hence the pxp + pxq + pxq + qxq probabilities. Or in table form
But certainly for a new mutation allele the q will be necessarily small, and thus it's relative proportion within the population will be overwhelmingly swamped by p in very large populations but not so much in very small populations. Enjoy Edited by RAZD, : tableby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
I don't think I have seen the Copelemurs on the charts before. Interesting. Is there a cladogram for this group? ... Sadly I don't have one, however I do have a link to the Gingrich paper where the grapic (afaik) originates (which is sort of like a cladogram bedded in the spatial\temporal matrix):
SYSTEMATICS, PHYLOGENY, AND EVOLUTION OF EARLY EOCENE ADAPIDAE (MAMMALIA, PRIMATES) IN NORTH AMERICA by Philip D. Gingerich and Elwyn L. Simons. see Page 32 for the graphic. There is also a long discussion of Copelemur and Pelycodus and their distinctions and relationships. Note that names used there differ from those used elsewhere (a not uncommon thing) and he gives reasons for these changes. The abstract is a good starting point:
quote: Note the discussion of the speciation event and divergence near the top of the graphic with one branch getting smaller while the other continues to get larger: "... Samples of species from successively higher stratigraphic horizons are progressively larger, until the second lineage of Pelycodus appears, after which one lineage begins to decrease in size while the other continues to increase. This is interpreted as a probable result of character divergence in body size to minimize competition between the two sympatric species. ... " -- this would be an example of the reinforcement selection you mentioned earlier. I also like where he says " ... The fossil record is now sufficiently complete that the boundary between Pelycodus and Notharctus is necessarily arbitrary. ... " as this is typical of what happens as more and more fossils are found. One can hope that a more complete picture\record of Copelemur and it's relationship to Pelycodus will also emerge with more fossils are found. This was written in 1977 after all. Scary thing, when doing a google on this one of the results was an EvC posting ... by me ...
Naw, Medical marijuana maybe? Painkillers for recent nasal basal cell carcinoma (non-malignant) surgery & skin graft (a friend says it looks like a nose piercing that went bad  ). ). Enjoy Edited by RAZD, : addedby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined: |
In this case, mating is not random but shows signs of inbreeding (the proportion of heterozygotes is reduced). We can estimate the inbreeding coefficient by F = (HO-HE)/HO where HO is heterozygotes observed and HE is heterozygotes expected. In this case F = 0.10 which means 10% of the population is autozygous. Evidence does not match expectation, it is the expectation (random mating) that is wrong.
Right, so we start with an allele frequency and determine the genotype. The population may then be subjected to selection which will remove an uneven proportion of alleles, ie. one allele will increase in frequency, the other will decrease. Selection models are kinda involved and I won't take time on it here, but let's say that the genotype aa is advantageous and the allele q increases from 0.10 to 0.15 because of selection. We now need to do the H-W calculation again to determine the genotypes of the next generation AA = 0.723, Aa = 0.255, aa = 0.022. So, an increase of 50% in allele frequency resulted in a 120% increase in 'aa' genotype frequency. Now if we consider {a} to have been a previously rare allele that is selected at a same normal frequency in the formation of the sub-population as it moved into a new habitat, ... and that it was beneficial there and increased 120% in a generation, ... but now the sub-population rejoins the parent population where {A} is still ~90%, and where {a} already existed as a selection neutral or mildly deleterious allele (hence the original rareness), ... this would not result in a significant rise of q in this recombined parent population, but one verging on stochastic variation within the large population from generation to generation. There would be virtually no difference in fitness of {aa} in this large population, no reason for selection to favor it this time around, no reason for it to expand in that population. So I don't see an existing, if rare, allele causing a significant change to the combined species population. There was a cause (selection, etc) that it was rare in the original population, and that cause would continue to operate. At best it could result in population subdivision if that new habitat continues to be used. This would act to keep {a} in the overall population, but still not become a major player in the overall population and not lead to takeover from the original population with a derived population. And even if taken to an extreme where the {a} allele achieves 90% frequency in the sub-population, when the populations rejoined the {aa} genotype could not be genetically incompatible with the parent population as it already exists there in small numbers (all things being equal and {aa} not being detrimental there). Certainly it cannot provide greater fitness in the original habitat as it already existed there. So I do not see this type of placid isolation and return changing things for the species population. But if we consider {a} to be derived by mutation in the small population then the picture changes. It enjoys faster selection in the small population and then when they rejoin the parent population (where q = 0) they would have an effect on the population. If the {aa} form is beneficial in the original habitat then it would continue to rise in frequency. It may or may not take over the whole population, depending on fitness. Enjoy Edited by RAZD, : clrtyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click)
|
|||||||||||||||||||||||||||||||||
|
RAZD Member (Idle past 1435 days) Posts: 20714 From: the other end of the sidewalk Joined:
|
See Extent of Mutational Capability, message 109 and Message 110, Message 113 and Message 123 -- recent evidence of four more recent Homo species interweaving behavior.
Also see A frog with a doubled genome -- interweaving evolution in frogs. So I would not be surprised to see more evidence of periods following population isolation, with subsequent isolated evolution of different traits, being occasionally interrupted by brief rejoining, with hybridizing and mixing of those traits, before reproductive incompatibility arises. The isolation can be due to geography\ecology and it can be due to different mating preferences in sub-populations. Certainly this means that speciation is not as cut and dried as was previously thought. If we consider that reproductive incompatibility is not a necessary aspect of the evolution of isolated sub-populations, that there is no selection pressure for that to occur, we end up with a lower bar for speciation -- mating preferences. The more the subspecies evolve different traits, the more members of each subspecies will prefer mates from their own population over the other, and then it becomes less likely that interbreeding occurs, but not ruled out. When the difference between such species reaches the same degree of differences that are used for phyletic (linear) speciation we are then able to call them different species. From Introduction to Evolution:
quote: This adds an amount of uncertainty and arbitrariness to divergent speciation (ibid):
quote: Perhaps instead we should say: " ... such that interbreeding becomes rare, then the formation of new species can be deemed to have occurred. ... " Enjoyby our ability to understand Rebel☮American☆Zen☯Deist ... to learn ... to think ... to live ... to laugh ... to share. Join the effort to solve medical problems, AIDS/HIV, Cancer and more with Team EvC! (click) |
|||||||||||||||||||||||||||||||||
|
|
Do Nothing Button
Copyright 2001-2023 by EvC Forum, All Rights Reserved
![]() ™ Version 4.2
™ Version 4.2
Innovative software from Qwixotic © 2024


 (1)
(1)